抗体,称为免疫球蛋白,通常是“Y”形,它们是由b细胞分泌的分子。抗体的基本功能是控制和阻止病原体并协助免疫反应。人的抗体可以分为5类(class),分别是IgA,IgD,IgE, IgG,IgM。以IgG为例,抗体是由两条相同的重链(H)和轻链(L)通过二硫键组装的四肽链结构。每条链具有恒定区(C)和可变区(V)。“Y”的尾部负责生物活性,例如与细胞结合。抗原结合区也称为互补位,是识别和结合抗原的部分。
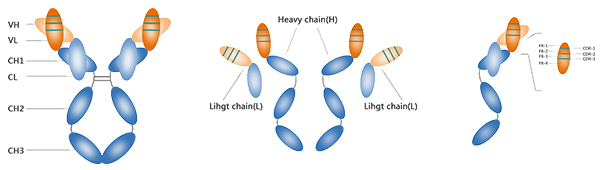
不同抗体的重链和轻链在靠近N端约110个氨基酸的序列变化很大,称为可变区(V区),重链和轻链的可变区分别称为VH和VL。重链和轻链在靠近C端的氨基酸序列相对稳定,称为恒定区(C区)。 重链和轻链的恒定区分别称为CH和CL。抗体的可变区用于识别和结合抗原,而恒定区则用于启动下游效应,比如抗体依赖性细胞介导的细胞毒作用(ADCC)。根据Ig轻链C区氨基酸组成和排列的不同,可将轻链分为κ和λ两型(type)。在λ型轻链C区内氨基酸组成又存在微小差异,由此将λ链分为4个亚型(subtype,λ1-λ4)。抗体的V区又可以细分为CDR(互补决定区)和FR(骨架区,FR1-FR4)。CDR区序列高度可变,而FR区序列则相对恒定。
抗体亚型(Antibody Subtype)
基于铰链区的大小,链间二硫键的位置和分子量的不同,人和鼠的IgG可以进一步细分为四种亚型。人的IgG可以分为IgG1(γ1),IgG2(γ2),IgG3(γ3),IgG4(γ4),按其在血清中的丰度(IgG1是最丰富的)的顺序命名。鼠的IgG可以分为IgG1,IgG2a,IgG2b,IgG3。在杂交瘤单抗制备完成后,需要对抗体亚型进行鉴定。南京j9九游会生物可以提供小鼠IgG1,IgG2a,IgG2b,IgG3及IgM,IgA,κ,λ链的鉴定,详情可联系。
IgG的功能
- IgG是血液和细胞外液中发现的主要抗体,可以控制身体组织的感染
- IgG是唯一可以穿过人体胎盘的免疫球蛋白,它主要负责在出生后的前几个月保护新生儿
- IgG是血液,淋巴液,脑脊液和腹膜液中的主要免疫球蛋白,是体液免疫反应的关键参与者
- IgG的Fc部分与吞噬细胞上的受体的结合是调理作用中的关键步骤。涂有IgG抗体颗粒的吞噬作用是细胞用于应对微生物的重要机制
- 由于其相对丰富和对抗原的优异特异性,IgG是用于免疫学研究和临床诊断的主要抗体
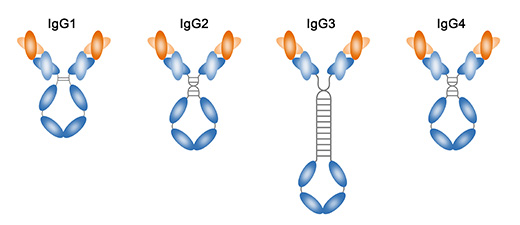
IgG1
IgG1占总IgG含量的60-65%左右。其主要负责胸腺介导的针对蛋白和多肽抗原的免疫应答。它还参与补体级联的调理和活化。IgG1同种型缺乏通常是低丙种球蛋白血症的征兆。
IgG2
IgG2占总IgG含量的20-25%左右。其主要针对碳水化合物/多糖抗原的免疫应答。在所有IgG同种型缺陷中,IgG2的缺乏,最常见的是引起婴儿的复发性气道/呼吸道感染。
IgG3
IgG3占总IgG含量的5-10%,在针对蛋白质或多肽抗原的免疫应答中起主要作用。
IgG4
IgG4通常占总IgG含量的4%以下,不与多糖结合。最新研究表明,在患有硬化性胰腺炎,胆管炎和间质性肺炎的患者中发现血清IgG4水平升高。目前有关IgG4的其他作用仍然未知。
IgG抗体亚型间的比较
| 对比项目 | IgG1(γ1) | IgG2(γ2) | IgG3(γ3) | IgG4(γ4) |
|---|---|---|---|---|
| 分子量(kDa) | 150 | 150 | 170 | 150 |
| 氨基酸隐藏区域 | 15 | 12 | 62 | 12 |
| H链间二硫键 | 2 | 4 | 11 | 2 |
| 半衰期(天) | 14-21 | 14-21 | 7 | 14-21 |
| 平均成人血清含量(g/L) | 6.98 | 3.8 | 0.51 | 0.56 |
| 相对丰度(%) | 60 | 32 | 4 | 4 |
IgG亚类之间氨基酸序列的细微差异也影响其生物学功能:
- IgG1,IgG3和IgG4容易穿过胎盘,但IgG2以非常低的效率穿过胎盘
- IgG3是最有效的补体激活剂,其次是IgG1; IgG2效率较低,IgG4根本不能激活补体
- IgG1和IgG3以高亲和力结合吞噬细胞上的Fc受体,从而介导调理作用。IgG4对Fc受体具有中等亲和力,IgG2具有极低的亲和力
抗体同种型(Antibody Isotype)
同一物种内所有个体共同具有的Ig抗原特异性结构。在哺乳动物中,有五种重链同种型:IgA(α),IgD(δ),IgE(ε),IgG(γ)和IgM(μ)。
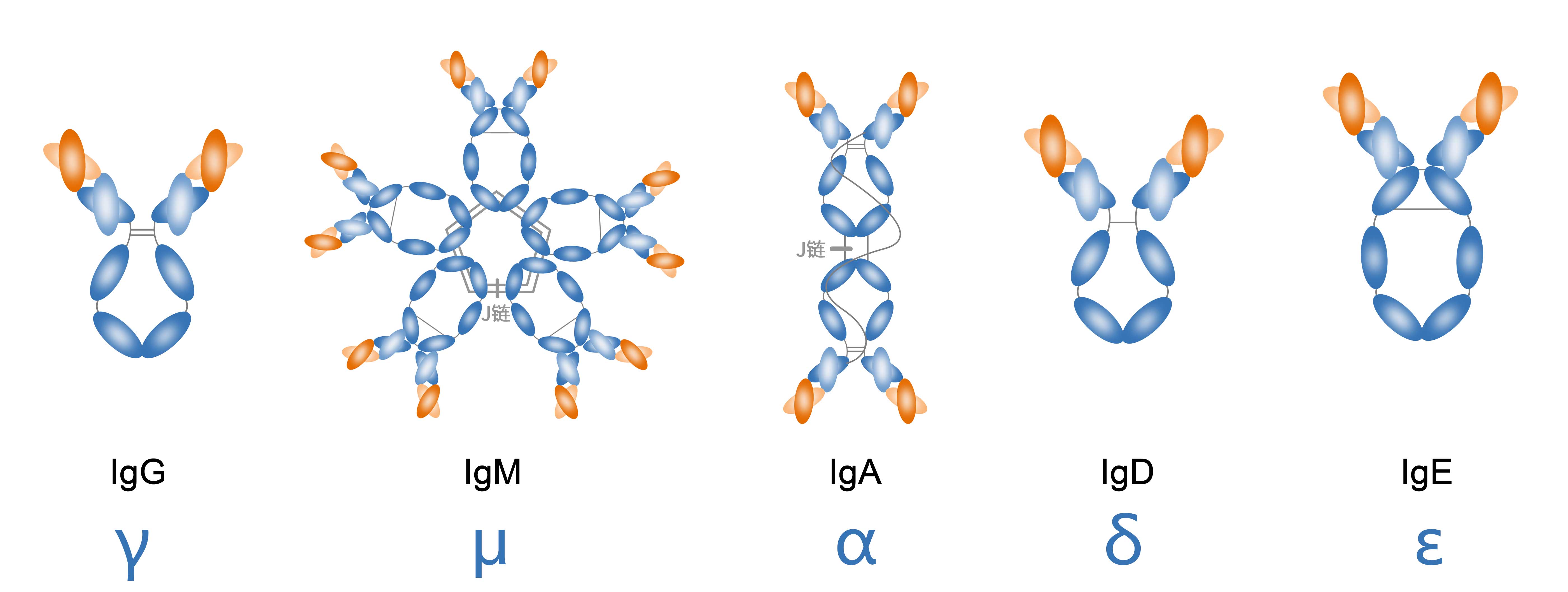
IgA(α)
IgA在呼吸道或肠道中分泌,充当粘膜免疫的主要介质,是抵御感染的第一道防线。它们在血清中是单体态,但在粘膜表面显示为二聚体。IgA抗体分为两个亚类,它们的铰链区大小不同。IgA1具有较长的铰链区,这增加了其对细菌蛋白酶的敏感性。因此,该亚类支配血清IgA,而IgA2主要存在于粘膜分泌物中。IgA的补体固定不是粘膜表面的主要效应机制,IgA受体在嗜中性粒细胞上表达,其可被激活以介导抗体依赖性细胞的细胞毒性。
IgD(δ)
IgD占免疫球蛋白库的比例不到1%,通常存在于B细胞的细胞膜上,其分子质量为150kDa。IgD同种型的表达水平与B细胞活化状态相关。目前,对于IgD的研究较少,人们对其在血清中的作用也知之甚少。
IgE(ε)
IgE通常存在于嗜碱性粒细胞和肥大细胞中,其分子质量约为190kDa。与IgM结构相似,IgE不含铰链区但具有两个额外的恒定结构域代替铰链区。IgE抗体与I型即时超敏反应有关。
IgG(γ)
IgG是血清中最丰富的抗体,占总免疫球蛋白库的70-85%,分子量约为150kDa,通常以单体形式存在。基于恒定区基因的结构差异和触发不同效应功能的能力将其分为4个亚类,IgG1、IgG2、IgG3和IgG4。彼此间具有高度的序列相似性(约90%相同),但是具有不同的半衰期,抗原结合特征和补体活化能力。IgG1是最丰富的IgG亚类,主导对蛋白质抗原的应答。对细菌荚膜多糖抗原的应答主要通过IgG2介导,它也是与聚糖抗原反应的主要抗体亚类。IgG3是促炎反应的有效激活剂,与其他IgG亚类相比,它具有最短的半衰期。IgG4是血清中最不丰富的IgG亚类,通常在反复接触相同抗原后或在持续感染期间产生。IgG的亚类中的IgG1、IgG3、IgG4可以穿过胎盘屏障,是唯一一种能够胎盘转运的免疫球蛋白,在新生儿抗感染免疫中起重要作用。
IgM(μ)
IgM是B细胞发育过程中表达的第一种免疫球蛋白,是初次免疫应答中的主要抗体,分子量约为900kDa。IgM能够以单体或五聚体的形式存在,与B细胞表面结合的是单体形式,在分泌形态中则是五聚体形式(通过J链连接)。五聚体结构使它们能够有效地结合抗原表位。由于IgM抗体在B细胞应答的早期表达,它们很少高度突变并具有广泛的抗原反应性,因此在不需要T细胞帮助的情况下提供对多种抗原的早期反应。